
腺垂体
最近審查:04.07.2025

所有iLive內容都經過醫學審查或事實檢查,以確保盡可能多的事實準確性。
我們有嚴格的採購指南,只鏈接到信譽良好的媒體網站,學術研究機構,並儘可能與醫學同行評審的研究相關聯。 請注意括號中的數字([1],[2]等)是這些研究的可點擊鏈接。
如果您認為我們的任何內容不准確,已過時或有疑問,請選擇它並按Ctrl + Enter。
脑垂体(学名:hypophysis,s.glandula pituitaria)位于蝶骨鞍区脑垂体窝内,脑垂体通过脑硬脑膜的突起与颅腔隔开,形成鞍膈。脑垂体通过鞍膈上的开口与间脑下丘脑的漏斗部相连。脑垂体横径为10-17毫米,前后径为5-15毫米,垂直径为5-10毫米。男性脑垂体重量约为0.5克,女性约为0.6克。脑垂体外部被一层囊膜包裹。
由于垂体由两个不同的胚胎发育而成,因此其可分为两个叶:前叶和后叶。腺垂体或前叶(腺垂体,s.lobus anterior)较大,占垂体总质量的70-80%。其密度高于后叶。前叶可分为远端部(远侧部),位于垂体窝前部;中间部(中间部),位于垂体窝与后叶的交界处;以及结节部(结节部),向上延伸并与下丘脑漏斗部相连。由于血管丰富,前叶呈淡黄色,略带红色。垂体前叶的实质由几种类型的腺细胞组成,腺细胞之间有窦状毛细血管。一半(50%)的腺垂体细胞是嗜色腺细胞,其细胞质中含有细颗粒,可被铬盐很好地染色。这些细胞是嗜酸性腺细胞(占所有腺垂体细胞的 40%)和嗜碱性腺细胞(10%)。嗜碱性腺细胞包括促性腺激素、促皮质素和促甲状腺激素内分泌细胞。嗜色腺细胞较小,具有较大的细胞核和少量的细胞质。这些细胞被认为是嗜色腺细胞的前体。另外 50% 的腺垂体细胞是嗜色腺细胞。
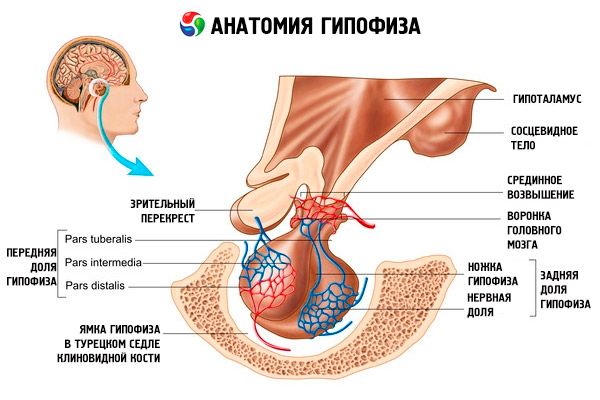
神经垂体,或称垂体后叶(neurohypophysis,s.lobus posterior),由位于垂体窝后部的神经叶(lobus nervosus)和位于腺垂体结节部后方的漏斗(infundibulum)组成。垂体后叶由神经胶质细胞(垂体细胞)、从下丘脑神经分泌核延伸至神经垂体的神经纤维以及神经分泌小体构成。
垂体通过神经纤维(通路)和血管,在功能上与间脑的下丘脑相连,下丘脑调节垂体的活动。垂体和下丘脑,连同它们之间的神经内分泌、血管和神经连接,通常被称为下丘脑-垂体系统。
垂体前叶和后叶的激素主要通过其他内分泌腺影响人体的诸多功能。在垂体前叶中,嗜酸性腺细胞(α细胞)产生生长激素,参与调节幼年生物体的生长发育过程。促皮质素内分泌细胞分泌促肾上腺皮质激素 (ACTH),这种激素刺激肾上腺分泌类固醇激素。促甲状腺素内分泌细胞分泌促甲状腺激素 (TSH),这种激素影响甲状腺的发育并激活其激素的产生。促性腺激素:促卵泡激素 (FSH)、促黄体生成素 (LH) 和催乳素——影响身体的性成熟,调节和刺激卵巢卵泡的发育、排卵、乳腺的生长和女性乳汁的分泌,以及男性精子的形成过程。这些激素由嗜碱性腺细胞(β细胞)产生。垂体的脂肪因子也分泌于此,影响体内脂肪的动员和利用。在前叶的中间部分,形成促黑素细胞激素,它控制着体内色素——黑色素——的形成。
下丘脑视上核和室旁核的神经分泌细胞产生加压素和催产素。这些激素沿着构成下丘脑-垂体束的轴突被输送到垂体后叶细胞。这些物质从垂体后叶进入血液。加压素具有血管收缩和抗利尿作用,因此也称为抗利尿激素(ADH)。催产素可以刺激子宫肌肉收缩,增加哺乳期乳腺的乳汁分泌,抑制黄体的发育和功能,并影响胃肠道平滑肌(非横纹肌)张力的变化。
垂体发育
垂体前叶由口湾背壁上皮发育而成,呈环状突起(Rathke囊)。该外胚层突起向未来第三脑室底部生长。第二脑泡下表面(未来第三脑室底部)向其方向生长出一个突起,漏斗部灰色结节和垂体后叶由此发育而成。
垂体的血管和神经
垂体上、下叶动脉从颈内动脉和大脑动脉环的血管通往垂体。垂体上、下叶动脉到达下丘脑的灰质核和漏斗部,在这里相互吻合,形成穿透脑组织的毛细血管——初级毛细血管网。从该网络的长环和短环中形成门静脉,通往垂体前叶。在垂体前叶的实质中,这些静脉分解成宽的窦状毛细血管,形成次级毛细血管网。垂体后叶的血液供应主要来自垂体下叶动脉。垂体上、下叶动脉之间存在长的动脉吻合口。来自次级毛细血管网络的静脉血通过静脉系统流出,流入脑硬脑膜的海绵窦和海绵间窦。
交感神经纤维随动脉进入垂体,参与垂体的神经支配。节后交感神经纤维从颈内动脉丛出发。此外,下丘脑核内的神经分泌细胞突起的众多末端位于垂体后叶。
垂体与年龄相关的特征
新生儿脑垂体的平均重量为0.12克。到10岁时,脑垂体的重量会增加一倍,到15岁时会增加两倍。到20岁时,脑垂体的重量达到最大值(530-560毫克),并且在随后的年龄段几乎保持不变。60岁以后,该内分泌腺的重量会略有下降。
 [ 7 ]、[ 8 ]、[ 9 ]、[ 10 ]、[ 11 ]、[ 12 ]、[ 13 ]、[ 14 ]、[ 15 ]
[ 7 ]、[ 8 ]、[ 9 ]、[ 10 ]、[ 11 ]、[ 12 ]、[ 13 ]、[ 14 ]、[ 15 ]
垂体激素
垂体和下丘脑之间紧密的解剖和功能联系,确保了体内神经和激素调节的统一性。这一复合体决定了整个内分泌系统的状态和功能。
垂体是主要的内分泌腺,它产生多种肽类激素,直接调节周围腺体的功能。它呈红灰色豆状结构,外覆纤维囊,重0.5-0.6克。其大小会因性别和年龄而略有不同。目前普遍接受的方法是将垂体分为两个叶,这两个叶在发育、结构和功能上有所不同:前远端垂体(腺垂体)和后垂体(神经垂体)。前垂体约占垂体总质量的70%,通常分为远端、漏斗部和中间部分;后垂体分为后部(或叶)和垂体柄。垂体位于蝶骨鞍区的垂体窝内,并通过垂体柄与大脑相连。前叶上部被视交叉和视束覆盖。垂体的血供非常丰富,由颈内动脉分支(垂体上、下垂体动脉)以及大脑动脉环的分支提供。垂体上动脉参与腺垂体的血供,垂体下动脉参与神经垂体的血供,并与下丘脑大细胞核轴突的神经分泌末梢相连。前者进入下丘脑正中隆起,并在那里分散形成毛细血管网络(初级毛细血管丛)。这些毛细血管(下丘脑内侧基底节的小神经分泌细胞的轴突末端与这些毛细血管相连)汇入门静脉,沿垂体柄下行至腺垂体实质,并再次在那里分成窦状毛细血管网络(次级毛细血管丛)。因此,血液先前流经下丘脑正中隆起,富含下丘脑促腺垂体激素(释放激素),之后进入腺垂体。
饱含腺垂体激素的血液从次级神经丛的众多毛细血管流出,通过静脉系统,流入硬脑膜的静脉窦,最终进入全身血流。因此,垂体的门静脉系统,其血流从下丘脑向下流动,是腺垂体营养功能的复杂神经体液调控机制的形态功能组成部分。
垂体由沿着垂体动脉走行的交感神经纤维支配。这些交感神经纤维起源于节后神经纤维,穿过颈内动脉丛,并与颈上神经节相连。下丘脑没有直接支配腺垂体。垂体后叶接受来自下丘脑神经分泌核的神经纤维。
腺垂体是一种组织结构非常复杂的器官。它有两种类型的腺细胞——嗜色腺细胞和嗜色腺细胞。嗜色腺细胞又分为嗜酸性腺细胞和嗜碱性腺细胞(垂体的详细组织学描述见本手册的相应章节)。然而,需要注意的是,由于腺垂体实质的多样性,构成腺垂体实质的腺细胞所分泌的激素在化学性质上存在一定差异,分泌细胞的精细结构也应与每种激素的生物合成特征相对应。但有时,在腺垂体中也可以观察到能够分泌多种激素的过渡型腺细胞。有证据表明,腺垂体的腺细胞类型并非总是由遗传决定的。
鞍膈下方是前叶的漏斗状部分。它包裹垂体柄,与灰质结节接触。腺垂体的这一部分以存在上皮细胞和丰富的血液供应为特征。它还具有激素活性。
垂体的中间部分由几层具有分泌活性的大型嗜碱性细胞组成。
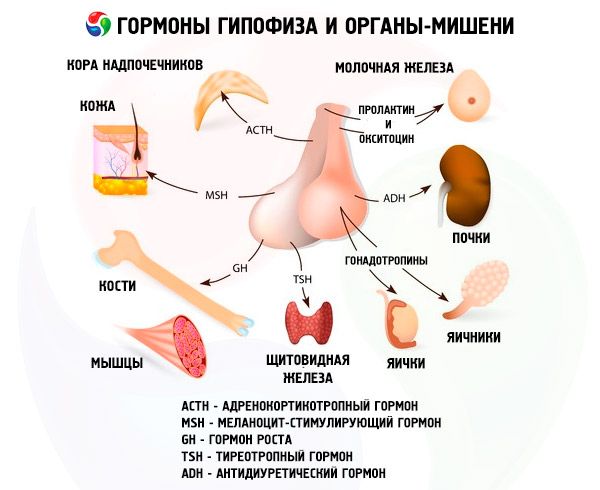
垂体通过其激素发挥多种功能。其前叶产生促肾上腺皮质激素 (ACTH)、促甲状腺激素 (TSH)、促卵泡激素 (FSH)、促黄体生成素 (LH)、促脂肪肝激素,以及生长激素 (STO) 和催乳素。中叶合成促黑素细胞激素 (MSH),后叶积累加压素和催产素。
促肾上腺皮质激素
垂体激素是一类蛋白质、肽类激素和糖蛋白。在垂体前叶的激素中,促肾上腺皮质激素(ACTH)的研究最为深入。它由嗜碱性粒细胞产生。其主要生理功能是刺激肾上腺皮质合成和分泌类固醇激素。ACTH还具有刺激黑素细胞和抗脂肪肝的活性。1953年,ACTH被分离纯化。之后,其化学结构被确定,在人类和许多哺乳动物中由39个氨基酸残基组成。ACTH没有物种特异性。目前,人们已经对ACTH本身及其分子的各种片段进行了化学合成,这些片段的活性高于天然激素。ACTH的结构包含两段肽链,其中一段确保ACTH与受体的识别和结合,另一段提供生物学效应。ACTH似乎通过激素与受体电荷的相互作用与ACTH受体结合。 ACTH的生物效应物的作用是由4-10分子(Met-Glu-His-Phen-Arg-Tri-Tri)的片段发挥的。
ACTH 的促黑素细胞活性源于其分子中存在一个 N 端区域,该区域由 13 个氨基酸残基组成,并重复了 α-促黑素细胞激素的结构。该区域还包含一个存在于其他垂体激素中的七肽,具有促肾上腺皮质激素、促黑素细胞活性和促脂肪生成活性。
ACTH 作用的关键环节被认为是在 cAMP 的参与下,细胞质中蛋白激酶的激活。磷酸化的蛋白激酶激活酯酶,酯酶将胆固醇酯转化为脂肪滴中的游离物质。细胞质中核糖体磷酸化产生的蛋白质刺激游离胆固醇与细胞色素 P-450 结合,并将其从脂质滴转移到线粒体,而线粒体中存在着所有确保胆固醇转化为皮质类固醇的酶。
[ 16 ]、[ 17 ]、[ 18 ]、[ 19 ]、[ 20 ]、[ 21 ]、[ 22 ]
促甲状腺激素
TSH(促甲状腺激素)是甲状腺发育和功能以及甲状腺激素合成和分泌过程的主要调节器。这种复杂的蛋白质——糖蛋白——由α亚基和β亚基组成。第一个亚基的结构与促黄体生成素的α亚基一致。而且,在不同动物物种中,其结构也基本一致。人类TSHβ亚基的氨基酸残基序列已被解析,由119个氨基酸残基组成。值得注意的是,人类和牛TSH的β亚基在许多方面相似。糖蛋白激素的生物学特性和生物活性取决于β亚基。它还确保激素与各种靶器官受体的相互作用。然而,大多数动物的β亚基只有与α亚基结合后才表现出特异性活性,而α亚基则充当激素的一种激活剂。后者以相同的概率诱导促黄体生成素、促卵泡生成和促甲状腺激素活性,这些活性由β亚基的特性决定。通过发现的相似性,我们可以得出结论,这些激素起源于一个共同的前身,β亚基也决定了这些激素的免疫学特性。有一种假设认为,α亚基保护β亚基免受蛋白水解酶的作用,并促进其从垂体运输到外周靶器官。
促性腺激素
促性腺激素以促黄体生成素 (LH) 和促卵泡激素 (FSH) 的形式存在于体内。这些激素的功能性目的通常仅限于确保男女个体的生殖过程。它们与促甲状腺激素 (TSH) 一样,是复杂的蛋白质——糖蛋白。FSH 诱导女性卵巢中卵泡的成熟,并刺激男性精子发生。LH 导致女性卵泡破裂,形成黄体,并刺激雌激素和孕激素的分泌。在男性中,LH 会加速间质组织的发育和雄激素的分泌。促性腺激素的作用相互依存,并同步发生。
女性促性腺激素的分泌动态在月经周期中发生变化,并已得到充分详细的研究。在月经周期的排卵前(卵泡期),LH(黄体生成素)含量较低,而FSH(卵泡刺激素)含量则升高。随着卵泡成熟,雌二醇分泌增加,从而促进垂体促性腺激素的分泌,并促进LH和FSH周期的发生,也就是说,性类固醇刺激促性腺激素的分泌。
目前,LH的结构已确定。与TSH类似,LH由两个亚基组成:a和b。不同动物物种的LH α亚基结构大致相同,与TSH α亚基的结构相对应。
LH β亚基的结构与 TSH β亚基的结构存在显著差异,尽管 LH β亚基具有四个相同的肽链片段,每个片段由 4-5 个氨基酸残基组成。在 TSH 中,这些片段位于 27-31、51-54、65-68 和 78-83 位。由于 LH 和 TSH 的 β亚基决定了激素的特定生物活性,因此可以假设 LH 和 TSH 结构中的同源片段应确保 β亚基与 α亚基的连接,而结构不同的片段则决定了激素生物活性的特异性。
天然LH对蛋白水解酶的作用很稳定,但β亚基很快被糜蛋白酶裂解,而a亚基则不易被酶水解,即起着保护作用,阻止糜蛋白酶接近肽键。
关于FSH的化学结构,研究人员尚未获得确切结果。与LH类似,FSH也由两个亚基组成,但FSH的β亚基与LH的β亚基不同。
催乳素
另一种激素——催乳素(催乳激素)在生殖过程中发挥着积极作用。催乳素在哺乳动物中的主要生理特性表现为刺激乳腺发育和泌乳,以及皮脂腺和内脏器官的生长。它促进类固醇对雄性第二性征的影响,刺激小鼠和大鼠黄体的分泌活动,并参与调节脂肪代谢。近年来,催乳素作为母性行为的调节剂受到了广泛关注;其多功能性可以通过其进化发展来解释。催乳素是一种古老的垂体激素,甚至在两栖动物中也存在。目前,一些哺乳动物物种中催乳素的结构已被完全破译。然而,直到最近,科学家们仍然对这种激素在人类中的存在表示怀疑。许多人认为它的功能是由生长激素实现的。目前,已有确凿证据表明催乳素存在于人体中,其结构也已部分解析。催乳素受体能够主动结合生长激素和胎盘催乳素,表明这三种激素具有单一的作用机制。
生长激素
生长激素(STH)的作用范围比催乳素更广。它与催乳素一样,由腺垂体的嗜酸性细胞产生。STH 能够刺激骨骼生长,激活蛋白质的生物合成,具有脂肪动员作用,并促进体型增长。此外,它还能协调代谢过程。
例如,当血糖水平下降时,垂体激素分泌量急剧增加,这证实了激素在后者中的参与。
这种人体激素的化学结构现已完全确定——由191个氨基酸残基组成。其一级结构与绒毛膜生长激素或胎盘催乳素相似。这些数据表明,尽管这两种激素在生物活性上存在差异,但它们在进化上具有显著的接近性。
有必要强调所讨论激素的高度物种特异性——例如,动物来源的STH在人类体内无活性。这既可以通过人类和动物STH受体之间的反应来解释,也可以通过激素本身的结构来解释。目前,正在进行研究,以识别STH复杂结构中表现出生物活性的活性中心。研究还研究了表现出其他特性的分子单个片段。例如,用胃蛋白酶水解人STH后,分离出一种由14个氨基酸残基组成、对应于分子31-44区的肽。它没有生长作用,但在脂肪代谢活性方面显著优于天然激素。与动物的类似激素不同,人类生长激素具有显著的催乳活性。
腺垂体合成多种具有脂肪动员作用的肽类和蛋白质物质,而促肾上腺皮质激素(ACTH)、促甲状腺激素(STH)、促甲状腺激素(TSH)等垂体促激素则具有促脂肪作用。近年来,β-和γ-促脂肪激素(LPG)的研究尤为突出。β-LPG的生物学特性研究最为深入;除了促脂肪作用外,它还具有刺激黑素细胞、促皮质素和降钙素血症的作用,并产生类似胰岛素的作用。
目前,绵羊LPG(90个氨基酸残基)、猪和牛的抗脂肪肝激素的一级结构已被解析。β-LPG中心区的结构在不同物种中相同,但该激素具有物种特异性。该区域的一个片段存在于α-MSH、β-MSH、促肾上腺皮质激素(ACTH)和β-LPG的结构中。这表明这些激素在进化过程中源自同一前体。γ-LPG的抗脂肪肝活性较β-LPG弱。
促黑素细胞激素
这种激素由脑垂体中叶合成,其生物学功能是刺激皮肤黑色素的生物合成,促进两栖动物皮肤中黑色素细胞的数量和大小增加。促黑素生成素(MSH)的这些特性被用于激素的生物学测试。该激素有两种类型:α-MSH 和 β-MSH。研究表明,α-MSH 没有物种特异性,在所有哺乳动物中都具有相同的化学结构。其分子是由 13 个氨基酸残基组成的肽链。相反,β-MSH 具有物种特异性,其结构在不同动物中有所不同。在大多数哺乳动物中,β-MSH 分子由 18 个氨基酸残基组成,只有在人类中,它的氨基酸末端才延长了 4 个氨基酸残基。值得注意的是,α-MSH 具有一定的促肾上腺皮质激素活性,其对动物和人类行为的影响现已得到证实。
催产素和加压素
下丘脑合成的加压素和催产素在垂体后叶积聚:加压素在视上核的神经元中积聚,催产素在室旁核中积聚。然后,它们被转移到垂体。需要强调的是,加压素激素的前体首先在下丘脑中合成。同时,1型和2型神经递质蛋白也在那里生成。前者与催产素结合,后者与加压素结合。这些复合物以神经分泌颗粒的形式在细胞质中沿着轴突迁移,到达垂体后叶,神经纤维在此终止于血管壁,颗粒内容物进入血液。加压素和催产素是第一种具有完整氨基酸序列的垂体激素。在化学结构上,它们是具有一个二硫键的九肽。
所讨论的激素会产生各种生物学效应:它们刺激水和盐通过细胞膜的运输,具有血管加压作用,增强分娩时子宫平滑肌的收缩,并增加乳腺的分泌。值得注意的是,加压素的抗利尿活性高于催产素,而催产素对子宫和乳腺的作用更强。加压素分泌的主要调节因素是水的消耗;在肾小管中,它与细胞质膜上的受体结合,随后激活其中的腺苷酸环化酶。该分子的不同部分负责将激素与受体结合并产生生物学效应。
垂体通过下丘脑与整个神经系统相连,将内分泌系统整合成一个功能整体,参与维持体内环境的稳定(体内平衡)。内分泌系统内的体内平衡调节基于垂体前叶与“靶”腺(甲状腺、肾上腺皮质、性腺)之间的反馈原理。“靶”腺分泌的激素过量会抑制相应的促激素,而其不足则会刺激相应的促激素的分泌和释放。下丘脑也属于反馈系统的一部分。对“靶”腺激素敏感的受体区就位于下丘脑中。下丘脑受体通过特异性结合血液循环中的激素,并根据激素浓度改变其反应,将其效应传递至相应的下丘脑中枢,这些中枢协调腺垂体的工作,释放下丘脑腺垂体促激素。因此,下丘脑应该被视为神经内分泌大脑。