
衣原體
最近審查:23.04.2024

所有iLive內容都經過醫學審查或事實檢查,以確保盡可能多的事實準確性。
我們有嚴格的採購指南,只鏈接到信譽良好的媒體網站,學術研究機構,並儘可能與醫學同行評審的研究相關聯。 請注意括號中的數字([1],[2]等)是這些研究的可點擊鏈接。
如果您認為我們的任何內容不准確,已過時或有疑問,請選擇它並按Ctrl + Enter。
衣原體-小革蘭氏陰性球菌寄生細菌屬於順序衣原體,衣原體家庭。目前,該家族包括兩種類型,其中在它們的抗原結構不同,細胞內的夾雜物和敏感性磺胺:衣原體(的沙眼衣原體衣原體(沙眼肺炎,:)鸚鵡熱衣原體)。
名稱“衣原體”(來自希臘語chtamys - 地幔)反映了微生物顆粒周圍存在膜。
所有類型的衣原體都具有共同的形態學特徵,常見的組抗原,分化的生殖週期。披衣菌被認為是已經失去合成ATP能力的革蘭氏陰性細菌。因此,它們是專性細胞內能量寄生蟲。
沙眼衣原體和肺炎衣原體歸因於人類無條件致病微生物,並且是人為染色體病的病原體。根據病原體類型和入口門(呼吸道,泌尿生殖系統),分離出呼吸道和泌尿生殖道衣原體病。
20在沙眼衣原體引起的多個實體,包括沙眼,結膜炎,腹股溝淋巴瘤,萊特爾綜合徵,泌尿生殖衣原體,沙眼衣原體感染引起的WHO估計第二中間等級描述的疾病,性傳播滴蟲感染後。全世界每年約有5000萬人登記註冊。
衣原體肺炎引起嚴重肺炎,上呼吸道疾病。有人猜測肺炎衣原體在動脈粥樣硬化和支氣管哮喘發病中的參與。
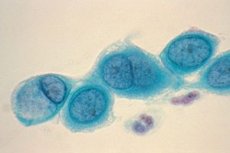
衣原體的形態和著色特性
衣原體是球形或卵形的小型革蘭氏陰性細菌。他們沒有鞭毛或膠囊。檢測衣原體的主要方法是Romanovsky-Giemsa染色。顏色的顏色取決於生命週期的階段:在細胞的藍色細胞質的背景中,初級環變成紫色,網狀體被染成藍色。
細胞壁的結構類似於革蘭氏陰性菌,儘管存在差異。它不含典型的肽聚醣:它完全含有N-乙酰胞壁酸。細胞壁包括包含LPS和蛋白質的外膜。儘管沒有肽聚醣,但衣原體的細胞壁是僵硬的。細胞的細胞質受到內部細胞質膜的限制。
衣原體外膜(HM)的分析表明,它包括LPS,外膜的主要蛋白質(MOMP)。以及富含半胱氨酸的蛋白質Omp1和Omp3,與NM的內表面相關。LPS和IOMP衣原體鸚鵡熱衣原體和沙眼衣原體不同於MHRP肺炎衣原體,位於細胞的外表面。分子量為90-100 kD的蛋白Omp Chitydia psittaci和肺炎衣原體也位於此處。
衣原體是多形的,這是由於它們繁殖的特性。唯一的(雙相)衣原體生命週期的特徵在於交替的兩種不同的形式存在 - 感染形式(原體 - ET)和自主形狀(網狀或初始細胞 - RT)。
微生物含有RNA和DNA。在RT中,RNA是DNA的4倍。在這些內容中是等價的。
網狀小體可以是橢圓形,半月形,雙極桿和球桿狀,大小為300-1000nm。網狀體不強制傳染性質,並且在分裂的情況下提供衣原體的繁殖。
初級卵形體,大小為250-500納米,具有感染性,可滲透進入發育週期的敏感細胞。它們具有緻密的外膜,這使得它們在細胞外環境中穩定。
衣原體培養
衣原體是專性寄生蟲,不能在人造營養培養基上繁殖,它們只能在活細胞中培養。它們是能量寄生蟲,因為它們不能獨立積累能量並使用宿主細胞的ATP。在HeLa細胞培養物McCoy中,在雞胚的卵黃囊中培養衣原體,敏感動物的身體在35℃的溫度下
 [3], [4], [5], [6], [7], [8], [9]
[3], [4], [5], [6], [7], [8], [9]
衣原體的抗原結構
衣原體具有三種類型的抗原:特異性抗原(在所有類型的衣原體中常見) - LPS; 信息特異性抗原(在所有類型的衣原體中不同) - 位於外膜中的蛋白質性質; 類型特異性(在血清型沙眼衣原體中不同) - 在微生物的細胞壁中產生的LPS; 蛋白質性質的變體特異性抗原。
血清型A,B,和C被稱為眼內,因為它們引起砂眼,血清型d,E,K,G,H,I,J,K(輝煌)是泌尿生殖道衣原體及其並發症,血清型L的病原體 - 激勵器性病淋巴肉芽腫病。呼吸病原體肺炎衣原體沙眼血清型4具有:TWAR,AR,RF,CWL。披衣菌psittaci有13個serovars。
[10], [11], [12], [13], [14], [15], [16], [17], [18]
衣原體的細胞取向
沙眼衣原體具有向泌尿生殖道上皮粘膜的趨向性,並且可以局部保留在其上或遍布組織表面。性病淋巴肉芽腫的病原體對淋巴組織具有向性。
肺炎衣原體在肺泡巨噬細胞,單核細胞和血管內皮細胞中增殖; 感染的全身傳播也是可能的。
鸚鵡熱衣原體在各種類型的細胞中引起感染,包括單核吞噬細胞。
衣原體發展的周期
沙眼衣原體的發育週期持續40-72小時,包括形態學和生物學性質不同的兩種存在形式。
在感染過程的第一階段,基本衣原體的吸附發生在敏感性宿主細胞的質膜上,並受到靜電力的作用。通過內吞作用將衣原體引入細胞。EG吸附在其上的質膜蛋白位點被陷入細胞質,形成吞噬泡。這個階段持續7-10小時。
進一步在6-8小時內,感染性元素體被重組為代謝活躍的非感染性,營養性,細胞內形式 - RT,其反複分裂。這些細胞內形式,即微菌落,被稱為衣原體包涵體。在發育的18-24小時內,它們位於由宿主細胞膜形成的細胞質囊泡中。包含物可以包含100至500個網狀衣原體。
在下一階段,在36-42小時內,發生成熟(形成中間體),並通過初級體內的分裂轉化網狀體。摧毀感染的細胞。基本的身體從它出現。在細胞外,初級細胞體在40-72小時後滲入新的宿主細胞,並開始衣原體發展的新循環。
除了該生殖週期之外,衣原體與宿主細胞的其他相互作用機制在不利條件下實現。這是吞噬體中衣原體的破壞,類似L型轉化和持久性。
轉化和持久形式的衣原體能夠轉化為原始(網狀)形式,隨後轉變為初級體。
在宿主細胞外,代謝功能被最小化。
衣原體致病性的因素
衣原體的粘附特性是由細胞外膜蛋白引起的,細胞外膜也具有抗細胞特性。另外,微生物細胞具有內毒素並產生外毒素。內毒素以LPS表示,在許多方面與LPS革蘭氏陰性細菌相似。不耐熱物質是外毒素,它們全部存在並在靜脈內給藥後導緻小鼠死亡。
在衣原體中,通過其將衣原體蛋白注射到宿主細胞的細胞質中的III型分泌系統的存在作為感染過程的組分發生。
熱休克蛋白(HSP)具有誘導自身免疫反應的性質。
[30], [31], [32], [33], [34], [35]
衣原體的生態和抗性
衣原體是一種非常常見的微生物。它們在超過200種動物,魚類,兩棲動物,軟體動物,節肢動物中被檢測到。高等植物中發現了形態相似的微生物。衣原體的主要宿主是人類,鳥類和哺乳動物。
致病菌在外界環境中不穩定,對高溫作用非常敏感,在乾燥時很快死亡。在50°C滅活30分鐘,在90°C - 1分鐘後。在室溫(18-20°C)下,病原體的感染活性在5-7天后下降。在37℃下,恆溫器中6小時的毒力下降80%。低溫(-20°С)促進病原體傳染性的長期保存。在紫外線照射的影響下,衣原體迅速死亡,與乙醚和70%乙醇接觸,10分鐘內2%賴氨醇,2%氯胺接觸。