
卵巢
Last reviewed: 04.07.2025

所有iLive內容都經過醫學審查或事實檢查,以確保盡可能多的事實準確性。
我們有嚴格的採購指南,只鏈接到信譽良好的媒體網站,學術研究機構,並儘可能與醫學同行評審的研究相關聯。 請注意括號中的數字([1],[2]等)是這些研究的可點擊鏈接。
如果您認為我們的任何內容不准確,已過時或有疑問,請選擇它並按Ctrl + Enter。
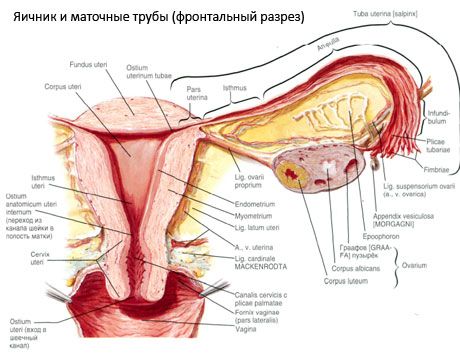
卵巢(ovary;希腊语:oophoron)是一对雌雄同体器官,是女性生殖腺,位于盆腔内子宫阔韧带后方。女性生殖细胞(卵子)在卵巢内发育成熟,并产生进入血液和淋巴液的女性性激素。卵巢呈卵圆形,前后略呈扁平状。卵巢呈粉红色。已生育女性的卵巢表面可见凹陷和疤痕——排卵和黄体发育的痕迹。卵巢重5-8克。卵巢尺寸:长2.5-5.5厘米,宽1.5-3.0厘米,厚达2厘米。卵巢有两个游离面:内侧面(facies medialis),面向盆腔,部分被输卵管覆盖;外侧面(facies lateralis),毗邻盆腔侧壁,略微凹陷,即卵巢窝。卵巢窝位于上方覆有腹膜的髂外血管与下方子宫动脉和闭孔动脉之间的夹角处。卵巢后方,相应侧输尿管自上而下经腹膜后行经。
卵巢表面延伸至凸起的自由(后)缘(margo liber),前方延伸至肠系膜缘(margo mesovaricus),并通过腹膜短褶(卵巢系膜)与子宫阔韧带后叶相连。卵巢前缘有一凹槽,即卵巢门(hilum ovarii),动脉和神经由此进入卵巢,静脉和淋巴管由此流出。卵巢也有两个末端:圆形的上部管状端(extremitas tubaria),朝向输卵管;以及下部子宫端(extremitas utenna),通过卵巢韧带(lig. ovarii proprium)与子宫相连。该韧带呈圆索状,粗约6毫米,从卵巢的子宫端延伸至子宫侧角,位于子宫阔韧带的两叶之间。卵巢的韧带装置还包括卵巢悬韧带(lig.suspensorium ovarii),它是腹膜的褶皱,从小骨盆壁延伸至卵巢,内部包含卵巢血管和纤维束。卵巢由短系膜(卵巢系膜)固定,系膜是腹膜的重复,从子宫阔韧带的后叶延伸至卵巢的肠系膜边缘。卵巢本身不被腹膜覆盖。输卵管最大的卵巢边缘附着于卵巢的管状末端。卵巢的形状取决于子宫的位置和大小(妊娠期间)。卵巢是小盆腔内活动性很强的器官。
 [
[ 卵巢的血管和神经
卵巢的血液供应由卵巢动脉和子宫动脉(aa. et vv. ovaricae et uterinae)提供。两条卵巢动脉(aa. dextra et sinistra ovaricae)均起源于肾动脉正下方的主动脉前表面;右侧卵巢动脉通常起源于主动脉,左侧卵巢动脉起源于肾动脉。每条卵巢动脉沿腰大肌前表面向下向外侧延伸,穿过前方的输尿管(向其发出分支)、髂外血管、边界线,进入盆腔,位于卵巢悬韧带内。卵巢动脉沿内侧方向穿过输卵管下方子宫阔韧带叶之间,向其发出分支,然后进入卵巢系膜;最终进入卵巢门。
卵巢动脉的分支与子宫动脉的卵巢分支广泛吻合。卵巢静脉的血液主要流出卵巢静脉丛,该丛位于卵巢门区域。血液从这里流出,分两个方向:子宫静脉和卵巢静脉。右侧卵巢静脉有瓣膜,汇入下腔静脉。左侧卵巢静脉没有瓣膜,汇入左肾静脉。
卵巢淋巴引流通过淋巴管进行,尤其在卵巢门区域最为丰富,此处卵巢下淋巴丛较为明显。淋巴随后沿着卵巢淋巴管排至主动脉旁淋巴结。
卵巢的神经支配
交感神经 - 由腹腔神经丛(太阳神经丛)、肠系膜上神经丛和腹下神经丛的节后纤维提供;副交感神经 - 由内脏骶神经提供。
卵巢的结构
卵巢表面覆盖一层单层生殖上皮。其下层为致密的结缔组织蛋白膜(白膜)。卵巢的结缔组织构成卵巢基质(卵巢基质),富含弹性纤维。卵巢实质分为外层和内层。内层位于卵巢中心,靠近卵巢门,称为卵巢髓质(卵巢髓质)。内层由疏松的结缔组织构成,包含众多血管、淋巴管和神经。卵巢的外层——卵巢皮质(卵巢皮质)——致密性更高。它含有大量的结缔组织,其中有成熟的初级卵泡(folliculi ovarici primarii)、次级(囊泡)卵泡(folliculi ovarici secundarii,s.vesiculosi),以及成熟卵泡、格拉夫卵泡(folliculi ovarici maturis)以及黄色和闭锁体。
每个卵泡都含有一个雌性生殖卵子,或称卵母细胞(ovocytus)。卵子直径可达150微米,呈圆形,含有细胞核和大量细胞质,除了细胞器外,还含有卵黄等蛋白质脂质内含物和糖原,这些都是卵子营养所必需的。卵子通常在排卵后12-24小时内耗尽其营养。如果未受精,卵子就会死亡。
人类卵子表面覆盖着两层膜。内层是细胞膜,也就是卵子的细胞质膜。细胞膜外是一层所谓的滤泡细胞,它们保护卵子,并发挥激素生成功能——分泌雌激素。
子宫、输卵管和卵巢的生理位置由连接腹膜、韧带和盆腔组织的悬吊、固定和支撑装置决定。悬吊装置由成对的结构组成,包括子宫圆韧带和阔韧带、子宫固有韧带和卵巢悬韧带。子宫阔韧带、卵巢固有韧带和卵巢悬韧带将子宫保持在中间位置。圆韧带将子宫底向前拉,并使其保持生理倾斜。
固定装置确保子宫位于小骨盆的中心位置,使其几乎不可能向两侧或前后移动。但由于韧带装置从子宫下部脱离,子宫可能向各个方向倾斜。固定装置包括位于骨盆疏松组织中的韧带,这些韧带从子宫下部延伸至骨盆的侧壁、前壁和后壁:骶骨韧带、主韧带、子宫膀胱韧带和膀胱耻骨韧带。
除卵巢系膜外,还有以下卵巢韧带:
- 卵巢悬韧带,以前称为漏斗骨盆韧带。它是腹膜的褶皱,卵巢的血管(a. et v. ovarica)、淋巴管和神经穿过它,延伸于骨盆外侧壁、腰筋膜(髂总动脉分为内外动脉的区域)和卵巢上端(输卵管)之间;
- 卵巢固有韧带穿过子宫阔韧带的两叶之间,靠近后叶,并将卵巢下端与子宫侧缘连接起来。卵巢固有韧带附着于子宫,位于输卵管起始部和圆韧带之间,圆韧带位于后者后上方。卵巢动脉后支(RR. Ovarii)穿过韧带的厚层,它是子宫动脉的终末分支;
- 阑尾-卵巢韧带:该韧带以腹膜皱襞的形式从阑尾系膜延伸至右侧卵巢或子宫阔韧带。该韧带不稳定,在1/2 - 1/3的女性中可见。
支撑装置由盆底的肌肉和筋膜表示,分为下层、中层和上层(内层)。
最强大的是上层(内层)肌肉,由提升肛门的成对肌肉组成。它由从尾骨向骨盆骨向三个方向(耻骨尾骨肌、髂尾骨肌和坐骨尾骨肌)散开的肌肉束组成。这一层肌肉也称为盆膈。
中层肌肉位于联合骨、耻骨和坐骨之间。中层肌肉——泌尿生殖膈——占据盆腔出口的前半部分,尿道和阴道由此穿过。在泌尿生殖膈的前部,各层肌肉之间是构成尿道外括约肌的肌束;在后部,是横向延伸的肌束——会阴深横肌。
盆底肌肉的下层(外层)由浅表肌肉组成,其形状类似于数字 8。这些肌肉包括球海绵体肌、坐骨海绵体肌、肛门外括约肌和浅表会阴横肌。
卵巢的个体发生
卵泡的生长和闭锁过程始于妊娠20周,到分娩时,女孩卵巢中残留的卵母细胞多达200万个。到月经初潮时,卵泡数量减少到30万个。在整个生殖寿命期间,只有不超过500个卵泡能够成熟并排卵。卵泡的早期生长不依赖于促卵泡激素(FSH)的刺激,生长受限,并且很快就会闭锁。目前认为,调节初级卵泡生长和闭锁的主要机制是局部自分泌/旁分泌肽,而非类固醇激素。卵泡的生长和闭锁过程被认为不受任何生理过程的干扰。该过程在所有年龄段都会持续,包括宫内期和更年期,并会因妊娠、排卵和无排卵而中断。触发卵泡生长及其在每个特定周期中的数量的机制尚不清楚。
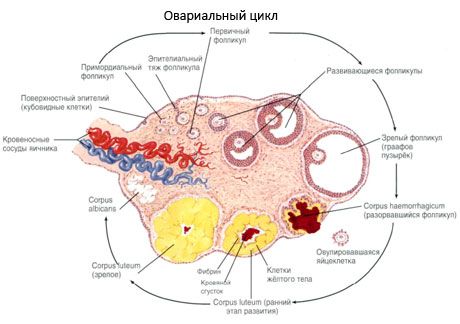
卵泡在发育过程中会经历几个阶段。原始生殖细胞起源于卵黄囊的内胚层和尿囊,并在妊娠5-6周时迁移至胚胎的生殖区域。由于快速的有丝分裂(从妊娠6-8周持续到妊娠16-20周),胚胎卵巢内形成了多达600万至700万个卵母细胞,这些卵母细胞被一层薄薄的颗粒细胞包裹。
腔前卵泡 - 卵母细胞被一层膜(透明带)包裹。卵母细胞周围的颗粒细胞开始增殖,其生长依赖于促性腺激素,并与雌激素水平相关。颗粒细胞是促卵泡激素(FSH)的靶细胞。在腔前卵泡阶段,颗粒细胞能够合成三类类固醇:主要诱导芳香化酶的活性,芳香化酶是将雄激素转化为雌二醇的主要酶。人们认为雌二醇能够增加自身受体的数量,从而独立于FSH对颗粒细胞发挥直接的促有丝分裂作用。它被认为是一种旁分泌因子,可以增强FSH的作用,包括激活芳香化过程。
卵泡开始生长后,FSH受体便出现在颗粒细胞膜上。FSH的减少或增加会导致其受体数量的变化。FSH的这种作用受生长因子的调节。FSH通过G蛋白(腺苷酸环化酶系统)发挥作用。虽然卵泡中的类固醇生成主要受FSH调控,但许多因素也参与了这一过程:离子通道、酪氨酸激酶受体、第二信使的磷脂酶系统。
雄激素在早期卵泡发育中的作用很复杂。颗粒细胞具有雄激素受体。这些受体不仅是FSH诱导的雌激素芳香化作用的底物,而且在低浓度下也能增强芳香化作用。当雄激素水平升高时,腔前颗粒细胞优先选择更简单的雄激素途径,而非雌激素芳香化途径,即通过5α还原酶转化为无法转化为雌激素的雄激素,从而抑制芳香化酶活性。该过程还会抑制FSH和LH受体的形成,从而阻止卵泡发育。
芳香化过程中,雄激素水平高的卵泡发生闭锁,卵泡的生长发育依赖于其将雄激素转化为雌激素的能力。
在FSH存在的情况下,卵泡液中的主要物质是雌激素。在FSH缺乏的情况下,卵泡液中则主要含有雄激素。通常情况下,LH在周期中期之前不存在于卵泡液中。LH水平一旦升高,颗粒细胞的有丝分裂活性就会降低,出现退行性改变,卵泡中的雄激素水平也会升高。卵泡液中的类固醇水平高于血浆中的类固醇水平,这反映了卵巢细胞(颗粒细胞和卵泡膜细胞)的功能活性。如果说FSH的唯一靶点是颗粒细胞,那么LH则有多个靶点——卵泡膜细胞、基质细胞、黄体细胞以及颗粒细胞。颗粒细胞和卵泡膜细胞都具有类固醇生成能力,但芳香化酶活性在颗粒细胞中占主导地位。
响应 LH,卵泡膜细胞产生雄激素,然后通过 FSH 诱导的芳香化,由颗粒细胞转化为雌激素。
随着卵泡的生长,卵泡膜细胞开始表达 LH 受体、P450 sec 和 3beta-羟基类固醇脱氢酶的基因,胰岛素样生长因子 (IGF-1) 与 LH 协同增加基因表达,但不会刺激类固醇生成。
卵巢类固醇生成始终依赖于促黄体生成素 (LH)。随着卵泡的生长,卵泡膜细胞会表达 P450c17 酶,该酶可将胆固醇转化为雄激素。颗粒细胞没有这种酶,依赖于卵泡膜细胞将雄激素转化为雌激素。与类固醇生成不同,卵泡生成依赖于促卵泡激素 (FSH)。随着卵泡生长和雌激素水平的升高,一种反馈机制被激活——FSH 生成受到抑制,这反过来又导致卵泡芳香化酶活性降低,最终导致卵泡因凋亡(程序性细胞死亡)而闭锁。
雌激素和促卵泡激素(FSH)的反馈机制会抑制已开始生长的卵泡发育,但不会抑制优势卵泡的发育。优势卵泡含有更多的FSH受体,支持颗粒细胞的增殖和雄激素芳香化为雌激素。此外,旁分泌和自分泌途径是窦状卵泡发育的重要协调者。
自分泌/旁分泌调节剂由肽类(抑制素、促活素、卵泡抑素)组成,这些肽类由颗粒细胞响应卵泡刺激素(FSH)合成并进入卵泡液。抑制素减少FSH的分泌;促活素刺激垂体释放FSH并增强FSH在卵巢中的作用;卵泡抑素可能通过与促活素结合来抑制FSH活性。排卵和黄体发育后,抑制素受促黄体生成素(LH)调控。
卵巢细胞的生长和分化受胰岛素样生长因子(IGE)的影响。IGF-1作用于颗粒细胞,引起环磷酸腺苷(cAMP)、孕酮、催产素、蛋白多糖和抑制素的增加。
IGF-1作用于卵泡膜细胞,导致雄激素分泌增加。卵泡膜细胞反过来又产生肿瘤坏死因子(TNF)和表皮生长因子(EGF),这些因子也受FSH调控。
EGF刺激颗粒细胞增殖,卵泡液中以IGF-2为主要生长因子,此外还含有IGF-1、TNF-a、TNF-3、EGF等。
卵巢功能的旁分泌和/或自分泌调节的破坏似乎在排卵障碍和多囊卵巢的发育中发挥作用。
随着窦卵泡的生长,卵泡液中的雌激素含量增加。在雌激素含量达到峰值时,颗粒细胞上会出现黄体生成素(LH)受体,颗粒细胞发生黄体化,孕酮生成增加。因此,在排卵前期,雌激素生成的增加会导致LH受体的出现,而LH又会导致颗粒细胞黄体化和孕酮生成。孕酮的增加会降低雌激素水平,这显然会导致在周期中期出现第二个FSH峰值。
排卵被认为发生在黄体生成素 (LH) 峰值后 10-12 小时,以及雌二醇峰值后 24-36 小时。LH 被认为能刺激卵母细胞减数分裂、颗粒细胞黄体化以及卵泡中孕酮和前列腺素的合成。
孕酮增强蛋白水解酶的活性,这些酶与前列腺素共同参与卵泡壁的破裂。孕酮诱导的FSH峰值通过将纤溶酶原转化为蛋白水解酶纤溶酶,使卵母细胞能够离开卵泡,并为黄体期的正常发育提供足够数量的LH受体。
排卵后3天内,颗粒细胞体积增大,并出现特征性空泡,其中充满色素——叶黄素。卵泡膜黄体细胞从卵泡膜和基质分化而来,成为黄体的一部分。在血管生成因子的影响下,穿透黄体的毛细血管迅速发育,随着血管化的改善,孕酮和雌激素的产生增加。黄体的类固醇生成活性和寿命取决于促黄体生成素(LH)的水平。黄体并非均质细胞结构。除了两种黄体细胞外,它还包含内皮细胞、巨噬细胞、成纤维细胞等。较大的黄体细胞产生肽(松弛素、催产素),并且比较小的黄体细胞更活跃于类固醇生成,芳香化酶活性更高,孕酮合成也更多。
孕酮峰值出现在黄体生成素 (LH) 峰值后第 8 天。值得注意的是,孕酮和雌二醇在黄体期周期性分泌,与 LH 的脉冲式分泌相关。随着黄体的形成,抑制素产生的控制权从 FSH 转移到 LH。在 LH 峰值之前,抑制素随着雌二醇的增加而增加,并在 LH 峰值之后继续增加,尽管雌激素水平会下降。虽然抑制素和雌二醇都是由颗粒细胞分泌的,但它们受不同的途径调控。黄体期末期抑制素的下降有助于下一个周期 FSH 的增加。
黄体减少非常快——在排卵后第9-11天。
退化的机制尚不清楚,与雌激素的黄体溶解作用或子宫内膜中观察到的受体相关机制无关。黄体产生的雌激素的作用还有另一种解释。已知雌激素是子宫内膜孕激素受体合成所必需的。黄体期雌激素可能是排卵后子宫内膜孕激素相关变化所必需的。由于雌激素水平不足导致孕激素受体发育不良,可能是不孕和早期流产的另一个机制,是黄体期缺陷的另一种形式。人们认为黄体的寿命是在排卵时确定的。如果妊娠期间得不到人绒毛膜促性腺激素的支持,黄体肯定会退化。因此,黄体退化会导致雌二醇、孕酮和抑制素水平下降。抑制素的降低会消除其对促卵泡激素(FSH)的抑制作用;雌二醇和孕酮的降低会使促性腺激素释放激素(GnRH)的分泌迅速恢复,并消除垂体的反馈机制。抑制素和雌二醇的降低,加上促性腺激素释放激素(GnRH)的升高,会导致FSH超过LH。FSH升高会导致卵泡生长,随后优势卵泡被选择,如果未受孕,则会开始新的周期。类固醇激素在生殖生物学和一般生理学中起着主导作用。它们决定人类的表型,影响心血管系统、骨代谢、皮肤、整体健康,并在妊娠中发挥关键作用。类固醇激素的作用反映了细胞内和遗传机制,这些机制对于将细胞外信号传递到细胞核并引起生理反应是必不可少的。
雌激素弥散地穿透细胞膜并与位于细胞核内的受体结合。受体-类固醇复合物随后与DNA结合。在靶细胞中,这些相互作用导致基因表达、蛋白质合成以及特定的细胞和组织功能。