
耶尔森氏菌
最近審查:08.07.2025

所有iLive內容都經過醫學審查或事實檢查,以確保盡可能多的事實準確性。
我們有嚴格的採購指南,只鏈接到信譽良好的媒體網站,學術研究機構,並儘可能與醫學同行評審的研究相關聯。 請注意括號中的數字([1],[2]等)是這些研究的可點擊鏈接。
如果您認為我們的任何內容不准確,已過時或有疑問,請選擇它並按Ctrl + Enter。
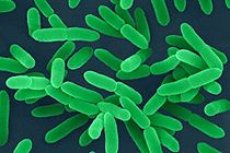
假结核耶尔森菌和小肠结肠炎耶尔森菌不属于特别危险的细菌,但它们在人类病理学中也起着重要作用。假结核耶尔森菌和小肠结肠炎耶尔森菌是多形性、不形成芽孢的革兰氏阴性杆菌,通常呈卵圆形,旧培养物中的细胞染色不均匀。从湿琼脂中提取的假结核菌可能具有双极染色,形成荚膜,但严重程度各不相同。与鼠疫耶尔森菌不同,这两种细菌都因有周生鞭毛而具有移动性。通过将其注射到半液体琼脂柱中检测移动性,但仅在 18-20°C 时才可检测,在 37°C 时则无移动性。耶尔森菌对培养基要求不高,在普通通用培养基中生长良好,并能在土壤和水中活跃繁殖。其生长最适温度为30°C,生长上限和下限分别为43°C和0-2°C,pH值范围为6.6-7.8。在Endo培养基中,24小时后,菌落直径为0.1-0.2毫米,呈圆形、凸起、有光泽、边缘光滑、无色(不发酵乳糖),几天后菌落大小为0.5-3毫米。R型假结核病菌落与鼠疫耶尔森菌落(中心有色素沉着,边缘呈扇贝状“蕾丝”状)几乎难以区分,但没有“碎玻璃”阶段。
所有三种类型的Yersinia抗原特性也有所不同。
假结核病原体根据O抗原分为8组(I-VIII),每组20个O因子抗原(1-20)。根据O和H抗原(ae),该种又分为13个血清型和亚血清型(la、lb、IIa、IIb、IIc、III、IVa、IVb、Va、Vb、VI、VII、VIII)。
小肠结肠炎耶尔森菌的特点是O抗原的异质性。该菌有34个血清型。大多数血清型适应特定动物物种或在环境中广泛存在。从人类中分离出的绝大多数菌株属于血清型03和09,较少见的是血清型06、08、05,非常罕见的是血清型01、02、010、011、013-017。
血清型I(LB),III和IV的菌株通常是从伪结核病患者中分离出来的。
在进化过程中,耶尔森氏菌获得了在两种环境中生存的需要——体外(腐生阶段)以及温血动物和人类体内(寄生阶段)。为了实现寄生阶段,耶尔森氏菌必须进入温血动物体内。食用在冰箱和蔬菜店中低温(4-12°C)储存的感染耶尔森氏菌的产品最常导致感染假结核病原体。在这些条件下,由于耶尔森氏菌的嗜冷性,它们可以在食物基质中繁殖和积累。这种感染方式的一个例子是1988年克拉斯诺达尔边疆区106名假结核病患者,其发病与食用感染了假结核耶尔森氏菌的卷心菜有关。土壤是其主要宿主。
在低温下,耶尔森氏菌具有较高的细胞和组织侵袭潜力,并能维持高毒力水平,但该病原体可能由于非特异性机制而通过任何黏膜穿透人体。野生和同人啮齿动物、家畜和农场动物也是耶尔森氏菌病的来源。人际传播是可能的。
已从175种哺乳动物、124种鸟类和7种鱼类中分离出假结核耶尔森氏菌菌株。受感染的啮齿动物、动物和人类会通过粪便和尿液排出病原体,污染水、植物和其他环境物质,人类通过这些物质被感染。因此,食物途径是假结核病和肠道耶尔森氏菌病原体传播的主要途径:感染是由于食用生的或未煮熟的食物(肉类、肉制品、牛奶、蔬菜、水果、绿叶蔬菜)造成的。这两种病原体不仅能够在植物上繁殖,还能在植物内部繁殖(例如生菜、豌豆、燕麦等)。
耶尔森菌引起的疾病具有多态性临床表现、胃肠道损害、全身蔓延倾向、败血症以及各器官和系统损害等特点。
小肠结肠炎耶尔森菌可导致人类胃肠炎,并损伤小肠壁。该病后常出现自身免疫性脊柱关节炎,例如赖特综合征和反应性关节炎。人们认为这些后果与小肠结肠炎耶尔森菌中存在超抗原有关。这些细菌的膜蛋白具有超抗原的特性。
远东地区的假结核病被描述为远东猩红热样热病。该病比西方地区的假结核病更为严重,其特点是过敏和毒性症状更为明显,尤其是在疾病的第二阶段。
耶尔森氏菌以及鼠疫病原体的致病特性不仅由染色体决定,还由质粒基因决定。它们的质粒与鼠疫耶尔森氏菌的质粒非常相似,编码VW抗原和外部蛋白(Yop)(与鼠疫耶尔森氏菌相同)以及其他毒力因子的合成。它们与鼠疫耶尔森氏菌具有与铁转运系统相关的共同基因簇。已证实,假结核耶尔森氏菌会合成一种耐热毒素,导致腹膜内感染的豚鼠死亡。病原体粘附和定植于肠粘膜的能力在假结核病的发病机制中起着重要作用。
耶尔森氏菌病的微生物诊断包括细菌学方法和血清学反应。细菌学方法中,将患者的待检物质(粪便、血液、咽部黏液)以及可疑产物或水接种于Endo、Ploskirev和Serov(指示和鉴别)培养基,并在37°C下培养48-72小时。将可疑菌落(在Endo和Ploskirev培养基上为小而无色的菌落,在Serov培养基上为两种不同形态的有色菌落)重新接种以获得纯培养物,根据生化特性进行鉴定,最后使用诊断性凝集血清进行分型。
假结核病和肠道耶尔森菌病的血清学诊断通常使用相应的诊断试剂进行详细的凝集反应(类似肥达氏反应),或使用抗原红细胞诊断试剂进行RPGA反应。抗体滴度达到1:400及以上为阳性。建议每隔几天进行一次双份血清反应。抗体滴度升高提示感染过程的特异性。